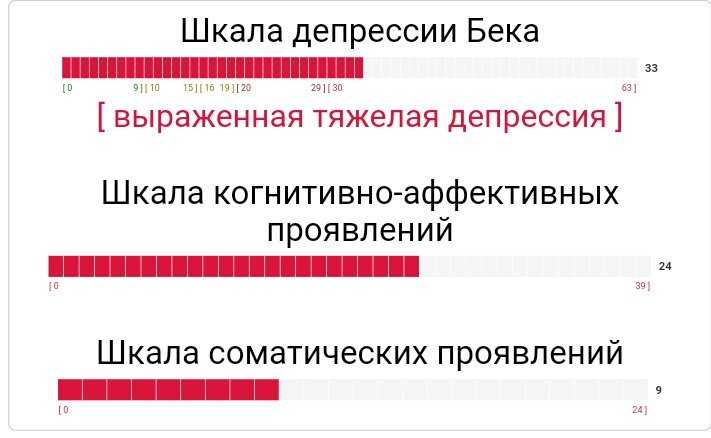
Тест на депрессию бека пройти тест: Шкала депрессии Э. Бека — Пройти онлайн тест
«Никогда» и «всегда»: ученые назвали 11 слов, которые часто произносят люди в депрессии — проверьте себя и близких | 45.ru
Все новостиЩит от «ЛГБТ-тьмы». Как и кого в России собираются наказывать за «нетрадиционные отношения»
Королева котельной. 26-летняя красотка с необычным именем стала сварщицей и мечтает открыть СТО
Дети сделают это легко: тест на возраст мозга — сможете найти на картинке все предметы?
«Вдруг им тоже потом скажут: «Мы вас туда не посылали?»» Ветеран Афгана — об участниках СВО на Украине
10 пищевых привычек, из-за которых мы толстеем и болеем
В Кургане в ряде домов отключили холодную воду
Бюджетные иномарки 2022 года: прицениваемся к недорогим альтернативам VW Polo и Hyundai Creta
«Наша арта долбит их, они — нас: в пинг-понг играем»: раненый военный — о службе, быте СВО и отряде «Ахмат»
Какие необычные случаи были в работе пожарных в Кургане прошлого века?
«Больше дождей и меньше снега»: какой погоды ждать от этой зимы в России
Из машин предпочитает Porsche. Что известно о 22-летней Карине, которая обокрала ювелирный магазин на 12 миллионов
Что известно о 22-летней Карине, которая обокрала ювелирный магазин на 12 миллионов
Пристроить можно и 5 тысяч: куда сегодня выгодно вкладывать деньги (если они у вас есть)
«Есть две категории — живые и мертвые». Узнали, почему мобилизованным отказывают в медкомиссии
Экс-мэра Екатеринбурга Евгения Ройзмана признали иноагентом
Кремль опроверг подготовку послания Путина о мобилизации всей страны: новости СВО за 25 ноября
Вместо машин и ресторанов — метро и сосиски: почему семья Тепляковых так бесит общественность — объясняет эксперт
Курганцы смогут высказать свои предложения по бюджету города на 2023 год
«Говорили, ничего не поможет»: женщина пять лет пыталась забеременеть и в итоге родила тройню (без ЭКО!)
Что Путин сказал матерям мобилизованных: онлайн
В Курганской области назвали размер тарифа на мусор в 2023 году
После какого возраста можно не думать о лишних килограммах. Ученые назвали идеальный вес для долголетия
На угольной игле: почему авария на «Листвяжной» и гибель 51 человека ничего не изменила и уже не изменит
Мобилизованные записали гневное видео о невыплате денег. После этого к ним приехали силовики
После этого к ним приехали силовики
Все говорят, что петь нельзя. Посмотрите «список запрещенных групп» — на этот раз в караоке-клубах
Жителей Курганской области предупредили об аномальных холодах
В частном секторе Кургана сгорел дом, погиб мужчина
Курганскому спортсмену привезли 200-килограммовый шар. Как он с ним тренируется?
В Кремле прокомментировали сообщения СМИ о готовящемся послании Путина о мобилизации страны
«Всё тупо для галочки»: доброволец рассказал о быте и подготовке к спецоперации в лагере для мобилизованных
Трагедия в Крымске: за что 66-летний пенсионер расстрелял трех человек
«Друзей там много, надеюсь увидимся»: зауралец записался добровольцем для участия в СВО
Житель Курганской области, осужденный за терроризм, пытался смягчить приговор
Запасаться гречкой и сахаром или скупать валюту в обменниках? Как на россиянах скажется милитаризация экономики
В Кургане при пожаре в общежитии по Автозаводской спасли 20 человек
В Курганской области у КАМАЗа отцепились колеса и влетели во встречные авто
В Курганской области многодетным мамам со знаками отличия пообещали выплаты
Мэрия Кургана ищет желающих ездить по новым маршрутам
Так пленку снимать не надо? Как на самом деле варить сосиски
Продлят ли маршрут № 42 до вокзала, как просили жители Утяка? Отвечает председатель гордумы Кургана
Все новости
Проверьте себя и своих близких — это несложно
 org/Person»>Фото: Дарья Селенская / Городские порталы
org/Person»>Фото: Дарья Селенская / Городские порталыПоделиться
Старшее поколение до сих пор часто не верит в такое заболевание, как депрессия. «Плохое настроение не болезнь, не придумывай», — говорят они. Но это не так. Это коварное и опасное заболевание, которое может сильно ухудшить качество жизни, а в крайних случаях — привести к печальным и непоправимым последствиям. Наши коллеги из Wday.ru разбирались, как распознать недуг у близкого, просто обратив внимание на его речь.
В отчете Всемирного экономического форума есть информация, что более 250 миллионов человек во всём мире мучаются от этого заболевания.
Депрессия — психическое расстройство, основными признаками которого являются сниженное — угнетенное, подавленное, тоскливое, тревожное, боязливое или безразличное — настроение и снижение или утрата способности получать удовольствие (ангедония).
Диагностировать депрессию в нашей сегодняшней реальности не так легко. Особенно если у человека непростая жизнь, ее часто путают с хронической усталостью. Но распознать болезнь важно, ведь, если она подтвердится, необходимо вмешаться специалисту. Вот несколько признаков депрессии:
Но распознать болезнь важно, ведь, если она подтвердится, необходимо вмешаться специалисту. Вот несколько признаков депрессии:
- подавленное настроение;
- усталость, лень, отсутствие энергии, заторможенность;
- потеря удовольствия от позитивных вещей;
- негативные мысли о себе — самокритика, самообвинения, мысли о своей ничтожности;
- физические признаки — нарушение сна, режима питания и веса.
При наличии таких признаков стоит пройти тест на депрессию «Шкала депрессии Бека» и провериться у специалиста. Если же депрессии нет, но жить тяжело и все ориентиры потеряны, то работать следует по .
При наличии таких признаков стоит пройти тест на депрессию «Шкала депрессии Бека» и провериться у специалиста. Если же депрессии нет, но жить тяжело и все ориентиры потеряны, то работать следует по этим четырем направлениям.
Исследование, опубликованное в научном журнале Clinical Psychological Science, помогает определить депрессию у близкого человека и вовремя помочь ему справиться с ней. Всё просто: надо его слушать и обращать особое внимание на слова, которые он использует в повседневной речи.
Всё просто: надо его слушать и обращать особое внимание на слова, которые он использует в повседневной речи.
Исследователи проанализировали 63 интернет-форума, всего в эксперименте участвовали более 64 000 человек. Они выявили закономерность — люди, которые страдали депрессией, часто использовали абсолютные выражения: «ничто», «никогда», «каждый» и «все».
— Выяснилось, что страдающие от депрессии используют много таких отрицательных прилагательных и наречий, как «одинокий», «печально», «грустно», «несчастный». Интересно то, что они используют гораздо больше личных местоимений в первом лице единственного числа: «я», «мне», «сам», что, возможно, отражает, насколько одинокими они чувствуют себя в этом мире, — рассказывает психолог Алиса Метелина.
Если вы долго пытаетесь вылечить аритмию, изжогу или даже заложенность носа, но ничего не помогает, возможно, дело не в сердце или ЖКТ. Под такими масками может скрываться депрессия, а причина недомоганий — химический сбой.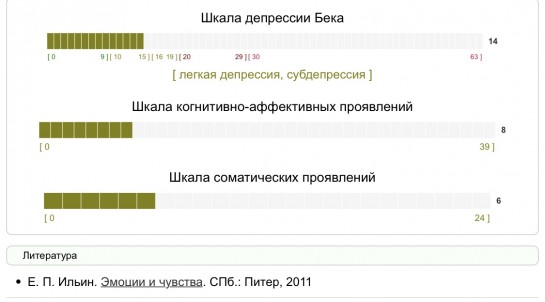 Как рассказала психотерапевт Татьяна Караваева, у маскированной депрессии те же причины и механизмы, что и у классической: нарушение обмена нейромедиаторов, а конкретно серотонина и норадреналина.
Как рассказала психотерапевт Татьяна Караваева, у маскированной депрессии те же причины и механизмы, что и у классической: нарушение обмена нейромедиаторов, а конкретно серотонина и норадреналина.
Татьяна Караваева — психотерапевт, руководитель отделения лечения пограничных психических расстройств и психотерапии НМИЦ психиатрии и неврологии им. Бехтерева, доктор медицинских наук.
— Это химический сбой, который возникает между нейронами головного мозга. И обычно это проявляется классической депрессией или, как говорят психиатры, триадой Крепелина. Она состоит из сниженного настроения, двигательной (моторной) и мыслительной (идеаторной) заторможенности. Это классика, — объясняет психотерапевт. — Но у некоторых людей развитие симптомов болезни идет не по обычному варианту, а по маскированному. Клинически такая депрессия будет проявляться не сниженным настроением, а соматическими симптомами, похожими на разные другие заболевания. Но эти симптомы никак не будут связаны с органическим повреждением, а будут результатом депрессии.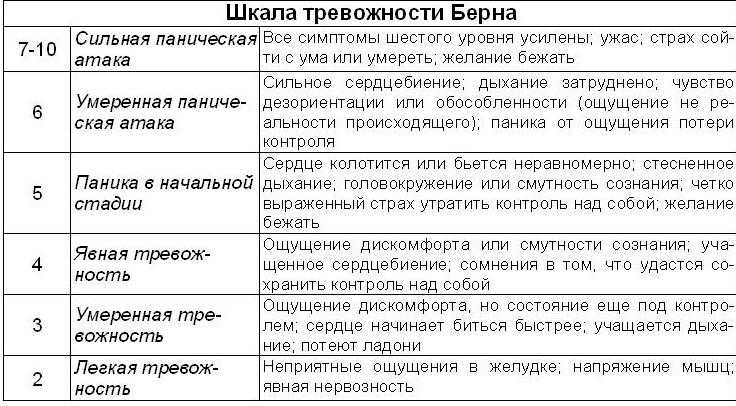
Вот и получается, что терапия, назначенная тем же кардиологом или гастроэнтерологом, таким пациентам не поможет.
— Эта терапия не воздействует на первопричину плохого самочувствия — химический сбой, связанный с серотонином и норадреналином. Как говорил известный психотерапевт Борис Дмитриевич Карвасарский, такие больные попадают в «большой медицинский лабиринт»: ходят от специалиста к специалисту, а к психотерапевтам приходят иногда уже спустя несколько лет от начала заболевания, когда формируются хронические формы, которые сложно лечить, — говорит специалист.
По словам Татьяны Караваевой, масок, за которыми может скрываться депрессия, на самом деле много и они разные. Врач рассказала о самых частых, которые встречаются в практике, и объяснила, как отличить депрессию от «настоящих» болезней.
- вредите себе каждый день: эксперты назвали 6 привычек несчастных людей — проверьте себя, вдруг вы тоже так делаете;
- эффект якоря: как работает самый популярный прием манипуляторов — с его помощью вас обманывают каждый день;
- привычка сутулиться: психологи объяснили, как опущенные плечи влияют на качество нашей жизни.


По теме
21 сентября 2022, 21:00
Схватились за сердце: как справиться с тревогой прямо сейчас — 4 простых приема и рейтинг лучших успокоительных14 сентября 2022, 22:00
Древние бы одобрили: эксперты рассказали, что значит желание укусить любимого человека на самом деле05 августа 2022, 15:00
Не говорить это ни в коем случае: правила общения с пожилыми родственниками с деменцией06 августа 2022, 17:00
Когда бокал вина за ужином становится проблемой? Как зарождается алкоголизм, разбираемся с наркологом03 ноября 2022, 15:00
Ни в одном глазу: врачи — о том, почему мы не можем уснуть, поможет ли мелатонин и какое время лучшее для сна
Мария Миронова
ДепрессияПсихологПсихотерапия
- ЛАЙК0
- СМЕХ0
- УДИВЛЕНИЕ0
- ГНЕВ0
- ПЕЧАЛЬ0
Увидели опечатку? Выделите фрагмент и нажмите Ctrl+Enter
КОММЕНТАРИИ0
Что я смогу, если авторизуюсь?
ПРАВИЛА КОММЕНТИРОВАНИЯ
0 / 1400Этот сайт защищен reCAPTCHA и Google. Применяются Политика конфиденциальности и Условия использования.
Применяются Политика конфиденциальности и Условия использования.
Новости СМИ2
Новости СМИ2
Тест на депрессию | шкала Бека
Выберите один из вариантов ответа: (1 из 21)
я чувствую себя хорошо мне плохо мне все время грустно, и я ничего не могу с собой поделать мне так скучно и грустно, что я не в силах больше терпеть
Выберите один из вариантов ответа: (2 из 21)
будущее не пугает меня я боюсь будущего меня ничто не радует мое будущее беспросветно
Выберите один из вариантов ответа: (3 из 21)
в жизни мне большей частью везло неудач и провалов было у меня больше, чем у кого-либо другого я ничего не добился в жизни я потерпел полное фиаско – как родитель, партнер, ребенок, на профессиональном уровне – словом, всюду
Выберите один из вариантов ответа: (4 из 21)
не могу сказать, что я неудовлетворен
как правило, я скучаю
что бы я ни делал, ничто меня не радует,
я как заведенная машина
меня не удовлетворяет абсолютно все
Выберите один из вариантов ответа: (5 из 21)
у меня нет ощущения, будто я кого-то обидел может, я и обидел кого-то, сам того не желая, но мне об этом ничего не известно у меня такое чувство, будто я всем приношу только несчастья я плохой человек, слишком часто я обижал других людей
Выберите один из вариантов ответа: (6 из 21)
я доволен собой иногда я чувствую себя несносным порой я испытываю комплекс неполноценности я совершенно никчемный человек
Выберите один из вариантов ответа: (7 из 21)
у меня не складывается впечатление, будто я совершил нечто такое, что заслуживает наказания я чувствую, что наказан или буду наказан за нечто такое, чему стал виной я знаю, что заслуживаю наказания я хочу, чтобы жизнь меня наказала
Выберите один из вариантов ответа: (8 из 21)
я никогда не разочаровывался в себе я много раз испытывал разочарование в самом себе я не люблю себя я себя ненавижу
Выберите один из вариантов ответа: (9 из 21)
я ничем не хуже других порой я допускаю ошибки просто ужасно, как мне не везет я сею вокруг себя одни несчастья
Выберите один из вариантов ответа: (10 из 21)
я люблю себя и не обижаю себя иногда я испытываю желание сделать решительный шаг, но не отваживаюсь лучше было бы вовсе не жить я подумываю о том, чтобы покончить жизнь самоубийством
Выберите один из вариантов ответа: (11 из 21)
у меня нет причин плакать бывает, что я и поплачу я плачу теперь постоянно, так что не могу выплакаться раньше я плакал, а теперь как-то не выходит, даже когда хочется
Выберите один из вариантов ответа: (12 из 21)
я спокоен я легко раздражаюсь я нахожусь в постоянном напряжении, как готовый взорваться паровой котел мне теперь все безразлично; то, что раньше раздражало меня, сейчас будто бы меня не касается
Выберите один из вариантов ответа: (13 из 21)
принятие решения не доставляет мне особых проблем иногда я откладываю решение на потом принимать решение для меня проблематично я вообще никогда ничего не решаю
Выберите один из вариантов ответа: (14 из 21)
мне не кажется, будто я выгляжу плохо или хуже, чем прежде меня волнует, что я неважно выгляжу я выгляжу плохо я безобразен, у меня просто отталкивающая внешность
Выберите один из вариантов ответа: (15 из 21)
совершить поступок – для меня не проблема мне приходится заставлять себя, чтобы сделать какой-либо важный в жизни шаг чтобы решиться на что-либо я должен очень много поработать над собой я вообще не способен что-либо реализовать
Выберите один из вариантов ответа: (16 из 21)
я сплю спокойно и хорошо высыпаюсь по утрам я просыпаюсь более утомленным, чем был до того, как заснул я просыпаюсь рано и чувствую себя невыспавшимся иногда я страдаю бессонницей, иногда просыпаюсь по несколько раз за ночь, в общей сложности я сплю не более пяти часов в суткиВыберите один из вариантов ответа: (17 из 21)
у меня сохранилась прежняя работоспособность я быстро устаю я чувствую себя уставшим, даже если почти ничего не делаю я настолько устал, что ничего не могу делать
Выберите один из вариантов ответа: (18 из 21)
аппетит у меня такой же, каким он был всегда у меня пропал аппетит аппетит у меня гораздо хуже, чем прежде у меня вообще нет аппетита
Выберите один из вариантов ответа: (19 из 21)
бывать на людях для меня так же приятно, как и раньше мне приходится заставлять себя встречаться с людьми у меня нет никакого желания бывать в обществе я нигде не бываю, люди не интересуют меня, меня вообще не волнует ничто постороннее
Выберите один из вариантов ответа: (20 из 21)
мои эротико-сексуальные интересы сохранились на прежнем уровне секс уже не интересует меня так, как прежде сейчас я мог бы спокойно обходиться без секса секс вообще не интересует меня, я совершенно потерял к нему влечение
Выберите один из вариантов ответа: (21 из 21)
я чувствую себя вполне здоровым и забочусь о своем здоровье так же, как и раньше у меня постоянно что-то болит со здоровьем дело обстоит серьезно, я все время об этом думаю мое физическое самочувствие ужасно, болячки просто изводят меня
Решение UnitedHealthcare по тесту на депрессию повышает перспективы Myriad
Статья из
Dive Brief
Опубликовано 2 августа 2019 г.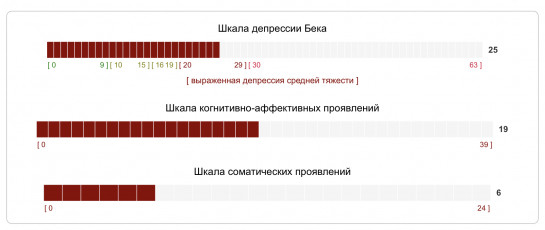
По
Ник Пол Тейлор Участник
Краткое описание погружения:
Гигант медицинского страхования UnitedHealthcare согласился покрыть стоимость теста на депрессию GeneSight компании Myriad Genetics, что подстегнуло усилия диагностической компании по увеличению продаж.
В отчете, опубликованном в четверг, Myriad сообщила, что решение распространяется на использование GeneSight у пациентов с большим депрессивным расстройством или тревогой, которые безуспешно пробовали по крайней мере одну предшествующую линию терапии.
- По оценкам аналитиков Cowen, решение о покрытии может добавить до 75 миллионов долларов к доходам Myriad в 2020 году. Аналитики понизили рейтинг акций Myriad в прошлом месяце, отчасти потому, что им не хватало «убежденности в перспективах более широкого покрытия GeneSight».

Dive Insight:
Решение UnitedHealthcare добавляет GeneSight в список мультигенных панелей, стоимость которых будет возмещена. С 1 октября страховщик будет возмещать расходы поставщикам услуг, заказавшим тест, при условии, что у пациента был диагностирован БДР или тревога, и он пробовал хотя бы одно лечение. Хотя CMS разрешает заказывать тест только психиатрам, UnitedHealthcare возместит запросы от всех поставщиков.
Обеспечение покрытия может повысить продажи продукта Бесчисленные надежды будут стимулировать рост. До этой новости Коуэн прогнозировал, что продажи GeneSight вырастут на 25% в следующем году. Теперь Myriad может превзойти этот прогноз.
«Наш прогноз доходов в настоящее время кажется консервативным, основываясь на положительном решении о покрытии [UnitedHealthcare] и возможном ознакомлении с другими частными плательщиками», — написали аналитики Cowen в примечании для инвесторов.
По оценкам аналитиков, решение UnitedHealthcare может увеличить продажи на сумму до 75 миллионов долларов в 2020 году.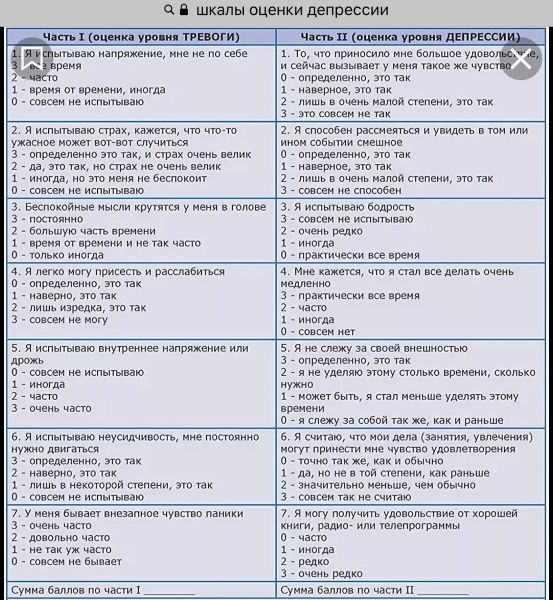 Myriad может увеличить свою выручку еще больше, если другие плательщики последуют примеру UnitedHealthcare или если решение о страховом покрытии приведет к тому, что рост объема превысит ожидания. Акции Myriad выросли на 54% после новостей о решении UnitedHealthcare.
Myriad может увеличить свою выручку еще больше, если другие плательщики последуют примеру UnitedHealthcare или если решение о страховом покрытии приведет к тому, что рост объема превысит ожидания. Акции Myriad выросли на 54% после новостей о решении UnitedHealthcare.
Этот ответ инвесторов отражает важность охвата GeneSight для планов роста Myriad. Myriad приобрела тест в результате поглощения Assurex Health за 225 миллионов долларов в 2016 году. В то время Myriad заявила, что рыночный потенциал теста превышает 4 миллиарда долларов.
Но GeneSight не смогла реализовать этот потенциал отчасти из-за отсутствия охвата плательщиков. За первые девять месяцев текущего финансового года Myriad получила доход в размере 82,9 млн долларов.
В 2017 году результаты ключевого клинического исследования с участием 1300 человек вызвали сомнения в том, что GeneSight когда-либо станет основным продуктом. Пациенты, оцененные с помощью теста, статистически показали не лучшие результаты по шкале депрессии, чем их коллеги, получавшие стандартное лечение.
UnitedHealthcare сослалась на исследование в своем решении о покрытии, отметив, что первичный результат «не отличался между двумя группами». Однако другие доказательства, в том числе тот факт, что «была значительная разница в ответе и ремиссии», заставили плательщика принять решение о покрытии теста.
- ФАРМАКОГЕНЕТИЧЕСКОЕ ТЕСТИРОВАНИЕ ЮнайтедЗдравоохранение
Рубрика: Политика и регулирование, Диагностика
Депрессивное опыление Lapageria Rosea Ruiz Et Pav.
-1
Депрессивное опыление Lapageria rosea Ruiz et Pav. (Philesiaceae) в фрагментированных тропических лесах умеренного пояса на юге Южной Америки КАРЛОС Э. ВАЛЬДИВИЯ*, ХАВЬЕР А. СИМОНЕТТИ и КАРОЛИНА А. ХЕНРИКЕС Отдел экологических наук, Факультет наук, Университет Чили, Касилья 653, Сантьяго, Чили ; *Автор для корреспонденции (e-mail: [email protected])
Ключевые слова: Фрагментация леса, Сохранение растений, Взаимодействие растений и животных, Аннотация винограда. Мы изучили опыление и репродуктивный успех в непрерывных и фрагментированных популяциях Lapageria rosea, самосовместимого растения, эндемичного для лесов умеренного пояса Чили. Численность опылителей, посещаемость, обилие цветков, объем и концентрация нектара, прорастание пыльцы и производство плодов и семян сравнивались между сплошным лесом площадью 145 га и четырьмя фрагментами леса площадью 6, 3, 3 и 1 га соответственно, окруженными зрелыми соснами. насаждения сосны лучистой. Обилие цветков было ниже в трех из четырех участков леса по сравнению со сплошным лесом. Объем нектара и концентрация сахара не различались между цветами в двух местах обитания. Опылители L. rosea, колибри Sephanoides sephaniodes и шмель Bombus dahlbomii, были менее многочисленны и реже посещали цветки L.
СИМОНЕТТИ и КАРОЛИНА А. ХЕНРИКЕС Отдел экологических наук, Факультет наук, Университет Чили, Касилья 653, Сантьяго, Чили ; *Автор для корреспонденции (e-mail: [email protected])
Ключевые слова: Фрагментация леса, Сохранение растений, Взаимодействие растений и животных, Аннотация винограда. Мы изучили опыление и репродуктивный успех в непрерывных и фрагментированных популяциях Lapageria rosea, самосовместимого растения, эндемичного для лесов умеренного пояса Чили. Численность опылителей, посещаемость, обилие цветков, объем и концентрация нектара, прорастание пыльцы и производство плодов и семян сравнивались между сплошным лесом площадью 145 га и четырьмя фрагментами леса площадью 6, 3, 3 и 1 га соответственно, окруженными зрелыми соснами. насаждения сосны лучистой. Обилие цветков было ниже в трех из четырех участков леса по сравнению со сплошным лесом. Объем нектара и концентрация сахара не различались между цветами в двух местах обитания. Опылители L. rosea, колибри Sephanoides sephaniodes и шмель Bombus dahlbomii, были менее многочисленны и реже посещали цветки L.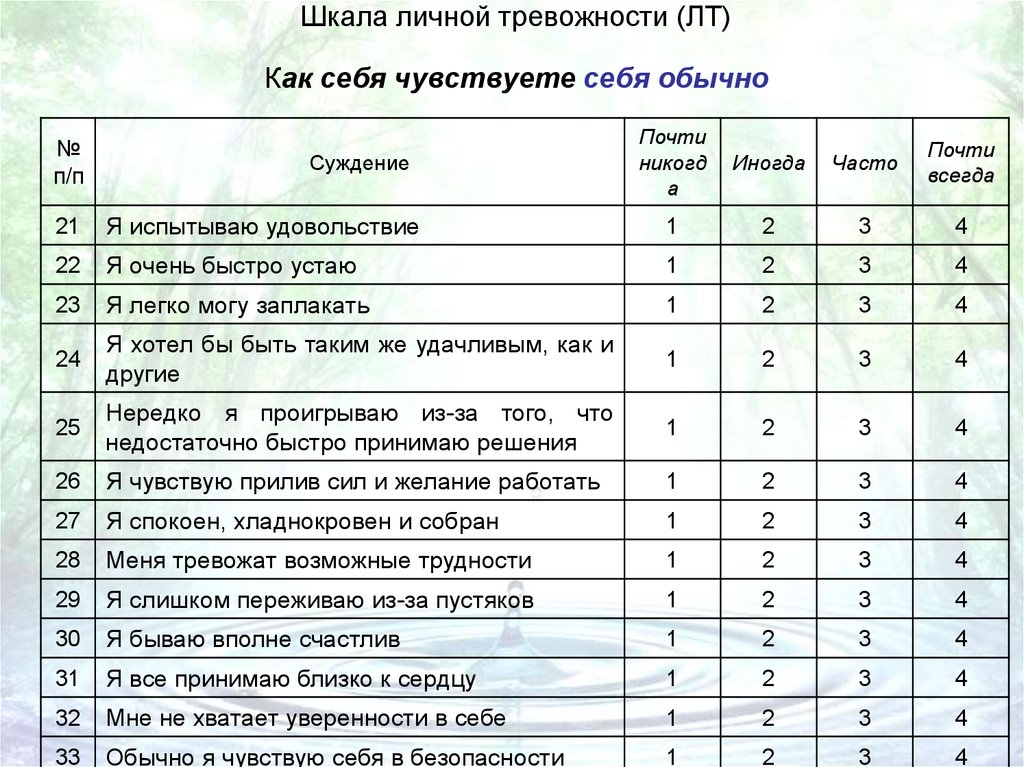
 87). Поскольку характеристики нектара зависят от условий окружающей среды (Zimmerman, 1988; Rathcke, 1992), фрагментация леса может также влиять на привлекательность растений для опылителей за счет изменений в скорости производства нектара и концентрации сахара. На самом деле концентрация сахара обычно увеличивается при низкой влажности за счет испарения воды в обмен на объем нектара. Колибри предпочитают концентрированный нектар до
30% концентрации сахара (Tamm and Gass 1986; Lo´pez-Calleja et al. 1997). Следовательно, высокая концентрация нектара может, в свою очередь, привести к более частому посещению растений опылителями (Southwick et al., 1981; Pleasants, 1983). Поскольку фрагментация среды обитания может снизить влажность воздуха (Капос, 1989; Матлак, 1993; Камарго и Капос, 1995), привлекательность цветов с точки зрения нектарных характеристик может быть выше в фрагментированных популяциях. Следовательно, фрагментация среды обитания может привести к одной из двух противоположных тенденций: снижению частоты посещения опылителей, поскольку небольшие и изолированные участки менее привлекательны; или увеличение количества посещений, потому что цветы в фрагментах могут производить более концентрированный нектар.
87). Поскольку характеристики нектара зависят от условий окружающей среды (Zimmerman, 1988; Rathcke, 1992), фрагментация леса может также влиять на привлекательность растений для опылителей за счет изменений в скорости производства нектара и концентрации сахара. На самом деле концентрация сахара обычно увеличивается при низкой влажности за счет испарения воды в обмен на объем нектара. Колибри предпочитают концентрированный нектар до
30% концентрации сахара (Tamm and Gass 1986; Lo´pez-Calleja et al. 1997). Следовательно, высокая концентрация нектара может, в свою очередь, привести к более частому посещению растений опылителями (Southwick et al., 1981; Pleasants, 1983). Поскольку фрагментация среды обитания может снизить влажность воздуха (Капос, 1989; Матлак, 1993; Камарго и Капос, 1995), привлекательность цветов с точки зрения нектарных характеристик может быть выше в фрагментированных популяциях. Следовательно, фрагментация среды обитания может привести к одной из двух противоположных тенденций: снижению частоты посещения опылителей, поскольку небольшие и изолированные участки менее привлекательны; или увеличение количества посещений, потому что цветы в фрагментах могут производить более концентрированный нектар. Более высокая концентрация нектарного сахара в цветках из фрагментов может компенсировать уменьшение размера цветочных пятен в фрагментах, что приводит к аналогичной частоте посещения растений опылителями и, как следствие, к аналогичной скорости образования семян. Сокращение размера популяции растений и усиление изоляции в фрагментированном ландшафте может вызвать увеличение спаривания среди близких родственников и, следовательно, вызвать депрессию инбридинга (Барретт и Кон 19).91; Эллстранд и Элам, 1993 г.; Янг и др. 1996). Пыльца, производимая особями с более высоким уровнем инбридинга, может иметь более низкую скорость прорастания, что влияет на производство семян (Johansson et al., 1998). Депрессия инбридинга также может выражаться в увеличении числа абортов плодов и семян и, следовательно, в снижении завязываемости семян (Ramsey and Vaughton 1996; Hauser and Siegismund 2000). В умеренных лесах центральной части Чили большое значение имеет опыление животными. Здесь посещается до 85% родов древесных растений и, вероятно, они опыляются животными (Айзен и др.
Более высокая концентрация нектарного сахара в цветках из фрагментов может компенсировать уменьшение размера цветочных пятен в фрагментах, что приводит к аналогичной частоте посещения растений опылителями и, как следствие, к аналогичной скорости образования семян. Сокращение размера популяции растений и усиление изоляции в фрагментированном ландшафте может вызвать увеличение спаривания среди близких родственников и, следовательно, вызвать депрессию инбридинга (Барретт и Кон 19).91; Эллстранд и Элам, 1993 г.; Янг и др. 1996). Пыльца, производимая особями с более высоким уровнем инбридинга, может иметь более низкую скорость прорастания, что влияет на производство семян (Johansson et al., 1998). Депрессия инбридинга также может выражаться в увеличении числа абортов плодов и семян и, следовательно, в снижении завязываемости семян (Ramsey and Vaughton 1996; Hauser and Siegismund 2000). В умеренных лесах центральной части Чили большое значение имеет опыление животными. Здесь посещается до 85% родов древесных растений и, вероятно, они опыляются животными (Айзен и др. , 2002). Леса в центральной части Чили сильно фрагментированы и характеризуются высоким уровнем эндемизма как животных, так и растений (Armesto et al. 19).96; Сан-Мартин и Доносо, 1996 г.; Бустаманте и Кастор, 1998). На сегодняшний день естественные леса в центральной части Чили сведены к многочисленным небольшим фрагментам, большинство из которых окружено плантациями экзотического дерева Pinus radiata L (Bustamante and Castor 1998). Таким образом, изучение влияния фрагментации местообитаний на опыление и его последствия для воспроизводства растений имеет большое значение для сохранения. В этой работе мы исследуем взаимодействие копиуэ (Lapageria rosea Ruiz et Pav. Philesiaceae) с их опылителями в фрагментированном ландшафте и то, как фрагментация лесов влияет на его репродуктивный успех. Лапагерия розовая — это лиана, эндемичная для этих лесов, и ее численность и плотность популяции в фрагментах леса ниже, чем в сплошном лесу (Henrı quez 2002). Опыление этого самосовместимого растения в основном зависит от двух видов животных: зеленоватой огненной кроны Sephanoides sephaniodes Lesson (Trochilidae) и гигантского шмеля Bombus dahlbomii Gu¨er.
, 2002). Леса в центральной части Чили сильно фрагментированы и характеризуются высоким уровнем эндемизма как животных, так и растений (Armesto et al. 19).96; Сан-Мартин и Доносо, 1996 г.; Бустаманте и Кастор, 1998). На сегодняшний день естественные леса в центральной части Чили сведены к многочисленным небольшим фрагментам, большинство из которых окружено плантациями экзотического дерева Pinus radiata L (Bustamante and Castor 1998). Таким образом, изучение влияния фрагментации местообитаний на опыление и его последствия для воспроизводства растений имеет большое значение для сохранения. В этой работе мы исследуем взаимодействие копиуэ (Lapageria rosea Ruiz et Pav. Philesiaceae) с их опылителями в фрагментированном ландшафте и то, как фрагментация лесов влияет на его репродуктивный успех. Лапагерия розовая — это лиана, эндемичная для этих лесов, и ее численность и плотность популяции в фрагментах леса ниже, чем в сплошном лесу (Henrı quez 2002). Опыление этого самосовместимого растения в основном зависит от двух видов животных: зеленоватой огненной кроны Sephanoides sephaniodes Lesson (Trochilidae) и гигантского шмеля Bombus dahlbomii Gu¨er.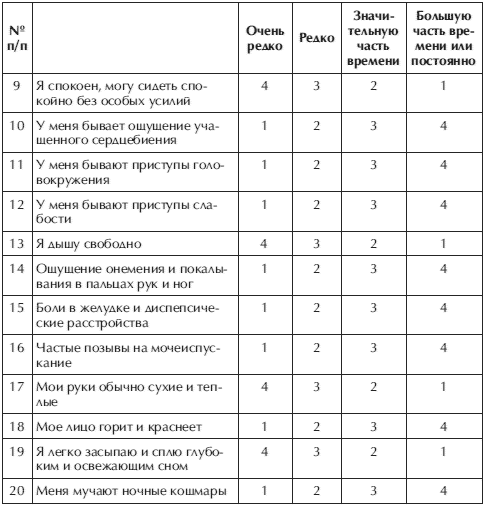 (Apidae) (Human˜a and Riveros 1994). Пик цветения приходится на осень, когда также цветет омела Tristerix corymbosus (L.) Kuijt, поскольку оба растения опыляются S. sephaniodes (Smith-Ramirez 1993; Henrıquez 2002). Если численность колибри и шмелей уменьшится в фрагментированном ландшафте, мы ожидаем более низкие показатели посещаемости L. rosea в более мелких лесных фрагментах. Следовательно, растения во фрагментах леса могут проявлять более низкую
успешность опыления, выраженная в меньшей доле оплодотворенных цветков, плодов и завязей семян. Более низкое качество переносимой пыльцы также может увеличить количество абортов плодов и семян. Однако, если снижение влажности воздуха увеличивает концентрацию нектара в более мелких фрагментах, мы ожидаем компенсаторного эффекта на посещаемость этих фрагментов и, возможно, нулевого эффекта на семенную продуктивность. Методы Место исследования Мы изучили фрагментированные и нефрагментированные популяции L. rosea в прибрежной зоне центральной части Чили (3559¢ S, 7241¢ W) в течение одного репродуктивного сезона, с мая 2001 г.
(Apidae) (Human˜a and Riveros 1994). Пик цветения приходится на осень, когда также цветет омела Tristerix corymbosus (L.) Kuijt, поскольку оба растения опыляются S. sephaniodes (Smith-Ramirez 1993; Henrıquez 2002). Если численность колибри и шмелей уменьшится в фрагментированном ландшафте, мы ожидаем более низкие показатели посещаемости L. rosea в более мелких лесных фрагментах. Следовательно, растения во фрагментах леса могут проявлять более низкую
успешность опыления, выраженная в меньшей доле оплодотворенных цветков, плодов и завязей семян. Более низкое качество переносимой пыльцы также может увеличить количество абортов плодов и семян. Однако, если снижение влажности воздуха увеличивает концентрацию нектара в более мелких фрагментах, мы ожидаем компенсаторного эффекта на посещаемость этих фрагментов и, возможно, нулевого эффекта на семенную продуктивность. Методы Место исследования Мы изучили фрагментированные и нефрагментированные популяции L. rosea в прибрежной зоне центральной части Чили (3559¢ S, 7241¢ W) в течение одного репродуктивного сезона, с мая 2001 г. по июнь 2002 г. Зона включала лес Маулино, богатый лиственными и вечнозелеными деревьями лес, в настоящее время сильно фрагментированный из-за лесного хозяйства (San Martı´n and Donoso 1996; Bustamante и Кастор 1998). В частности, мы работали в сплошном лесу, в национальном заповеднике Лос-Кеулес (охраняемая территория площадью 145 га, вложенная в лесную колею площадью 600 га), и в четырех фрагментах естественного леса небольшого размера (от 1 до 6 га), все их окружают плантации Pinus radiata. Фрагменты леса менее влажны, чем сплошной лес, круглый год (Henrı quez 2002). Плотность цветков, объем и концентрация нектара Мы оценивали густоту цветков как количество цветов на участке площадью 25 м2. Участки были случайным образом распределены по участкам. Плотность цветков оценивали в пик сезона цветения L. rosea в мае 2002 г. (Henrıquez 2002). Мы измеряли объем нектара в спелых цветках в мешках в течение 24 ч (n = 35 в сплошном лесу, n = 35 в лесных фрагментах), многократно вводя микрокапиллярную трубку объемом 60 мкл в шесть нектарников каждого цветка до тех пор, пока не перестанет извлекаться нектар.
по июнь 2002 г. Зона включала лес Маулино, богатый лиственными и вечнозелеными деревьями лес, в настоящее время сильно фрагментированный из-за лесного хозяйства (San Martı´n and Donoso 1996; Bustamante и Кастор 1998). В частности, мы работали в сплошном лесу, в национальном заповеднике Лос-Кеулес (охраняемая территория площадью 145 га, вложенная в лесную колею площадью 600 га), и в четырех фрагментах естественного леса небольшого размера (от 1 до 6 га), все их окружают плантации Pinus radiata. Фрагменты леса менее влажны, чем сплошной лес, круглый год (Henrı quez 2002). Плотность цветков, объем и концентрация нектара Мы оценивали густоту цветков как количество цветов на участке площадью 25 м2. Участки были случайным образом распределены по участкам. Плотность цветков оценивали в пик сезона цветения L. rosea в мае 2002 г. (Henrıquez 2002). Мы измеряли объем нектара в спелых цветках в мешках в течение 24 ч (n = 35 в сплошном лесу, n = 35 в лесных фрагментах), многократно вводя микрокапиллярную трубку объемом 60 мкл в шесть нектарников каждого цветка до тех пор, пока не перестанет извлекаться нектар. Мешки были изготовлены из тюлевой сетки для уменьшения возможных перепадов температуры и влажности внутри цветка и окружающей среды, что позволяет избежать ошибок при оценке объема нектара и концентрации сахара. Измерения проводились каждые 2 часа в течение дня, с 8:30 до 18:30, в период с марта по май 2002 г. Одновременно с измерением объема нектара мы оценивали концентрацию сахара в нектаре, выраженную в % мас./мас. . Измерения проводились ручным рефрактометром с температурной компенсацией. Численность опылителей Численность опылителей оценивали как количество особей S. sephaniodes и B. dahlbomii, зарегистрированных визуальными учетами на 48 стационарных станциях в сплошном лесу и на 48 во фрагментах. Каждая станция имела радиус 10 м, а расстояние между ними составляло примерно 30 м. Наблюдения проводились через 10 мин.
интервалы с 8:00 до 12:00. Это время соответствует пику активности обоих опылителей (Смит-Рамирез и Арместо 19).98)
Частота посещений и пыльцевая нагрузка Мы оценили частоту посещений случайно выбранных цветков опылителями в течение 10-минутных периодов наблюдения в течение дня с 8:00 до 17:00 в период с марта по июнь 2001 г.
Мешки были изготовлены из тюлевой сетки для уменьшения возможных перепадов температуры и влажности внутри цветка и окружающей среды, что позволяет избежать ошибок при оценке объема нектара и концентрации сахара. Измерения проводились каждые 2 часа в течение дня, с 8:30 до 18:30, в период с марта по май 2002 г. Одновременно с измерением объема нектара мы оценивали концентрацию сахара в нектаре, выраженную в % мас./мас. . Измерения проводились ручным рефрактометром с температурной компенсацией. Численность опылителей Численность опылителей оценивали как количество особей S. sephaniodes и B. dahlbomii, зарегистрированных визуальными учетами на 48 стационарных станциях в сплошном лесу и на 48 во фрагментах. Каждая станция имела радиус 10 м, а расстояние между ними составляло примерно 30 м. Наблюдения проводились через 10 мин.
интервалы с 8:00 до 12:00. Это время соответствует пику активности обоих опылителей (Смит-Рамирез и Арместо 19).98)
Частота посещений и пыльцевая нагрузка Мы оценили частоту посещений случайно выбранных цветков опылителями в течение 10-минутных периодов наблюдения в течение дня с 8:00 до 17:00 в период с марта по июнь 2001 г. (Kearns and Inouye 1993). . Отбор проб проводили в течение двадцати 10-минутных периодов в сутки и 4 дня в месяц на сплошном лесу и два по 3 дня на участках леса. Общее количество наблюдений соответствовало 253 периодам в сплошном лесу и 153 периодам в фрагментах леса. Чтобы проверить, эффективно ли S. sephaniodes и B. dahlbomii переносят пыльцевые зерна L. rosea, мы подсчитали количество пыльцевых зерен L. rosea среди пыльцы других видов, переносимых на телах животных. Извлечение пыльцы колибри (n = 3) производили, отлавливая птиц паутинными сетями и накладывая на голову и клюв каждой птицы липкую прозрачную липкую ленту. Затем клейкие ленты анализировали непосредственно под микроскопом. Для получения пыльцевых зерен шмелей мы отлавливали животных энтомологическим сачком (n = 17), которых промывали 5 мл 50%-ного раствора этанола (Kearns, Inouye 19).93). Наконец, мы подсчитали количество пыльцевых зерен L. rosea в шести аликвотах с помощью гемацитометра под микроскопом. Пыльцевую нагрузку на шмеля оценивали как среднее количество из шести аликвот, экстраполированных на весь объем образца.
(Kearns and Inouye 1993). . Отбор проб проводили в течение двадцати 10-минутных периодов в сутки и 4 дня в месяц на сплошном лесу и два по 3 дня на участках леса. Общее количество наблюдений соответствовало 253 периодам в сплошном лесу и 153 периодам в фрагментах леса. Чтобы проверить, эффективно ли S. sephaniodes и B. dahlbomii переносят пыльцевые зерна L. rosea, мы подсчитали количество пыльцевых зерен L. rosea среди пыльцы других видов, переносимых на телах животных. Извлечение пыльцы колибри (n = 3) производили, отлавливая птиц паутинными сетями и накладывая на голову и клюв каждой птицы липкую прозрачную липкую ленту. Затем клейкие ленты анализировали непосредственно под микроскопом. Для получения пыльцевых зерен шмелей мы отлавливали животных энтомологическим сачком (n = 17), которых промывали 5 мл 50%-ного раствора этанола (Kearns, Inouye 19).93). Наконец, мы подсчитали количество пыльцевых зерен L. rosea в шести аликвотах с помощью гемацитометра под микроскопом. Пыльцевую нагрузку на шмеля оценивали как среднее количество из шести аликвот, экстраполированных на весь объем образца. Качество пыльцы Оценивали качество пыльцы, получая пыльцу с цветков из сплошного леса и фрагментов (один цветок на растение, n = 34 в сплошном лесу и n = 37 в фрагментах). Прорастание пыльцы оценивали как процент пыльцы, проросшей in vitro в среде Брюбейкера-Квака при 20% сахарозы (Kearns and Inouye 19).93). Пыльцевые зерна считали проросшими, когда сформировалась пыльцевая трубка. Оптимальная концентрация раствора сахарозы была предварительно определена в ходе испытаний на прорастание при шести различных концентрациях сахарозы (1, 2,5, 5, 10, 20 и 30% масс./масс.). В каждом образце подсчитывали количество проросших пыльцевых зерен в шести аликвотах с помощью гемацитометра под микроскопом. Количество пыльцевых зерен, проросших на один цветок, оценивали как среднее из шести аликвот.
Завязь семян и плодов Для оценки завязываемости семян и плодов мы пометили 46 зрелых цветков в сплошном лесу и 44 во фрагментах леса, по одному цветку на растение, следующие за ними.
в течение года. Мы оценили долю оплодотворенных цветков, недоразвитых плодов и недоношенных плодов и, наконец, долю цветков, успешно завязывающих плоды на каждом участке.
Качество пыльцы Оценивали качество пыльцы, получая пыльцу с цветков из сплошного леса и фрагментов (один цветок на растение, n = 34 в сплошном лесу и n = 37 в фрагментах). Прорастание пыльцы оценивали как процент пыльцы, проросшей in vitro в среде Брюбейкера-Квака при 20% сахарозы (Kearns and Inouye 19).93). Пыльцевые зерна считали проросшими, когда сформировалась пыльцевая трубка. Оптимальная концентрация раствора сахарозы была предварительно определена в ходе испытаний на прорастание при шести различных концентрациях сахарозы (1, 2,5, 5, 10, 20 и 30% масс./масс.). В каждом образце подсчитывали количество проросших пыльцевых зерен в шести аликвотах с помощью гемацитометра под микроскопом. Количество пыльцевых зерен, проросших на один цветок, оценивали как среднее из шести аликвот.
Завязь семян и плодов Для оценки завязываемости семян и плодов мы пометили 46 зрелых цветков в сплошном лесу и 44 во фрагментах леса, по одному цветку на растение, следующие за ними.
в течение года. Мы оценили долю оплодотворенных цветков, недоразвитых плодов и недоношенных плодов и, наконец, долю цветков, успешно завязывающих плоды на каждом участке. Мы также оценили количество семян в плоде.
Результаты Плотность цветков и нектаропродукция Обилие цветков на фрагментах леса было в 1,4 раза ниже, чем в сплошном лесу (0,6 ± 0,2 цветка/25 м2 на фрагментах и 0,9± 0,2 цветка/25 м2 в сплошном лесу, среднее значение ± SE Критерий Манна-Уитни, U = 4036, p = 0,002). В пределах фрагментов три из четырех лесных фрагментов имели обилие цветов ниже, чем сплошной лес (разброс: 0,16 ± 0,08–1,04 ± 0,31 цветков/25 м2 в фрагментах). Следовательно, общее предложение цветков опылителям ниже во фрагментах, чем в сплошном лесу. Объем нектара и концентрация на цветок были одинаковыми между фрагментами и сплошным лесом, колеблясь от 22 мкл до 38 мкл в течение дня в сплошном лесу и фрагментах (рис. 1, повторное измерение ANOVA, F1,5 = 0,69)., р = 0,63). Концентрация сахарозы в нектаре также была одинаковой между сплошным лесом и фрагментами (t = 1,51, df = 127, p = 0,14) и составила 17,01 ± 4,72%. мас./мас. в фрагментах леса и 18,33 ± 4,75% мас./мас. в сплошном лесу (среднее ± SE).
Мы также оценили количество семян в плоде.
Результаты Плотность цветков и нектаропродукция Обилие цветков на фрагментах леса было в 1,4 раза ниже, чем в сплошном лесу (0,6 ± 0,2 цветка/25 м2 на фрагментах и 0,9± 0,2 цветка/25 м2 в сплошном лесу, среднее значение ± SE Критерий Манна-Уитни, U = 4036, p = 0,002). В пределах фрагментов три из четырех лесных фрагментов имели обилие цветов ниже, чем сплошной лес (разброс: 0,16 ± 0,08–1,04 ± 0,31 цветков/25 м2 в фрагментах). Следовательно, общее предложение цветков опылителям ниже во фрагментах, чем в сплошном лесу. Объем нектара и концентрация на цветок были одинаковыми между фрагментами и сплошным лесом, колеблясь от 22 мкл до 38 мкл в течение дня в сплошном лесу и фрагментах (рис. 1, повторное измерение ANOVA, F1,5 = 0,69)., р = 0,63). Концентрация сахарозы в нектаре также была одинаковой между сплошным лесом и фрагментами (t = 1,51, df = 127, p = 0,14) и составила 17,01 ± 4,72%. мас./мас. в фрагментах леса и 18,33 ± 4,75% мас./мас. в сплошном лесу (среднее ± SE). Численность опылителей В целом численность опылителей была примерно в 1,4 раза ниже на фрагментах леса, чем в сплошном лесу (критерий Манна-Уитни, U = 603,
Рис. 1. Объем нектара Lapageria rosea в сплошном и фрагментированном лесу (среднее ± SE).
р 0,001. Рисунок 3а). Ни один из фрагментов не представил количество посещений на цветок выше, чем сплошной лес (диапазон: 0,06 ± 0,04–0,24 ± 0,06 посещения/цветок/10 мин). Основное различие в частоте посещений давал B. dahlbomii. Фактически, относительная важность посещений B. dahlbomii в сплошном лесу составила 57,5% от общего числа посещений (рис. 3с), а на фрагментах леса — 35,0% от общего числа посещений. У Sephanoides sephaniodes было на 60% меньше посещений во фрагментах, чем в сплошном лесу (критерий Манна-Уитни, U = 539).4, р = 0,03, рис. 3б), в то время как у B. dahlbomii было на 80% меньше посещений во фрагментах, чем в сплошном лесу (критерий Манна-Уитни, U = 4960, р = 0,003, рис. 3в). Две из трех особей S. sephaniodes и пятнадцать из семнадцати особей B.
Численность опылителей В целом численность опылителей была примерно в 1,4 раза ниже на фрагментах леса, чем в сплошном лесу (критерий Манна-Уитни, U = 603,
Рис. 1. Объем нектара Lapageria rosea в сплошном и фрагментированном лесу (среднее ± SE).
р 0,001. Рисунок 3а). Ни один из фрагментов не представил количество посещений на цветок выше, чем сплошной лес (диапазон: 0,06 ± 0,04–0,24 ± 0,06 посещения/цветок/10 мин). Основное различие в частоте посещений давал B. dahlbomii. Фактически, относительная важность посещений B. dahlbomii в сплошном лесу составила 57,5% от общего числа посещений (рис. 3с), а на фрагментах леса — 35,0% от общего числа посещений. У Sephanoides sephaniodes было на 60% меньше посещений во фрагментах, чем в сплошном лесу (критерий Манна-Уитни, U = 539).4, р = 0,03, рис. 3б), в то время как у B. dahlbomii было на 80% меньше посещений во фрагментах, чем в сплошном лесу (критерий Манна-Уитни, U = 4960, р = 0,003, рис. 3в). Две из трех особей S. sephaniodes и пятнадцать из семнадцати особей B.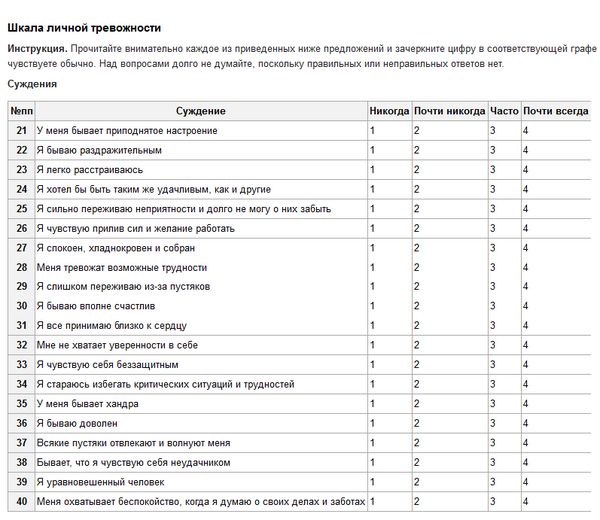 dahlbomii несли пыльцу L. rosea. Восемьдесят пять процентов пыльцевых зерен, переносимых S. sephaniodes (29,7 ± 25,7 пыльцевых зерен/птица, среднее ± SE), и 96 % пыльцевых зерен, переносимых B. dahlbomii (294,2 ± 107,4 пыльцевых зерен/шмель), приходится на пыльцу Л. розовая.
Всхожесть пыльцы Всхожесть пыльцы, собранной с цветков лесных фрагментов, была примерно на 50 % ниже, чем пыльцы цветков, произрастающих в сплошном лесу (19).0,8% ± 13,7 в фрагментах и 33,4% ± 11,9 в сплошном лесу; среднее ± SE. Критерий Манна-Уитни, U = 278, p 0,05. Таблица 1). Однако количество семян на плод было примерно на 12% ниже во фрагментах, чем в сплошном лесу (80,7 ± 3,4 семян/плод в сплошном лесу и 69,27 ± 4,5 семян/плод во фрагментах леса, t = 2,018; df = 128, p = 0,05). ). Количество семян на плод колебалось от 51,2 ± 13,11 до 76,91 ± 6,66 семян/плод на фрагментах леса.
Обсуждение Lapageria rosea, несмотря на то, что она самосовместима, нуждается в опылителях для завязывания семян, поскольку является автогамной (Human˜a and Riveros 19).
dahlbomii несли пыльцу L. rosea. Восемьдесят пять процентов пыльцевых зерен, переносимых S. sephaniodes (29,7 ± 25,7 пыльцевых зерен/птица, среднее ± SE), и 96 % пыльцевых зерен, переносимых B. dahlbomii (294,2 ± 107,4 пыльцевых зерен/шмель), приходится на пыльцу Л. розовая.
Всхожесть пыльцы Всхожесть пыльцы, собранной с цветков лесных фрагментов, была примерно на 50 % ниже, чем пыльцы цветков, произрастающих в сплошном лесу (19).0,8% ± 13,7 в фрагментах и 33,4% ± 11,9 в сплошном лесу; среднее ± SE. Критерий Манна-Уитни, U = 278, p 0,05. Таблица 1). Однако количество семян на плод было примерно на 12% ниже во фрагментах, чем в сплошном лесу (80,7 ± 3,4 семян/плод в сплошном лесу и 69,27 ± 4,5 семян/плод во фрагментах леса, t = 2,018; df = 128, p = 0,05). ). Количество семян на плод колебалось от 51,2 ± 13,11 до 76,91 ± 6,66 семян/плод на фрагментах леса.
Обсуждение Lapageria rosea, несмотря на то, что она самосовместима, нуждается в опылителях для завязывания семян, поскольку является автогамной (Human˜a and Riveros 19). 94). Следовательно, растения, произрастающие во фрагментированных лесах Маулино, могут оказаться в демографическом водовороте (sensu Soule, 1986). Сниженное опыление приводит к угнетенной приспособленности по сравнению с особями, живущими в сплошных лесах. Сценарий, с которым сталкивается L. rosea, может быть репрезентативным для других видов, опыляемых колибри и шмелями в находящихся под угрозой южных тропических лесах умеренного пояса Южной Америки. Каждая особь L. rosea производит меньше семян в результате снижения опыления и снижения способности пыльцевых зерен к прорастанию. В целом, это снижение репродуктивного успеха может, в свою очередь, привести размер популяции к демографическому вихрю, который может привести к исчезновению. Ни плотность нектара, ни плотность цветков не объясняют снижение уровня опыления L. rosea во фрагментах. Несмотря на более низкую влажность воздуха во фрагментах, одинаковое количество нектара с той же концентрацией сахара производят цветы, независимо от того, произрастают ли они в лесных остатках или сплошном лесу.
94). Следовательно, растения, произрастающие во фрагментированных лесах Маулино, могут оказаться в демографическом водовороте (sensu Soule, 1986). Сниженное опыление приводит к угнетенной приспособленности по сравнению с особями, живущими в сплошных лесах. Сценарий, с которым сталкивается L. rosea, может быть репрезентативным для других видов, опыляемых колибри и шмелями в находящихся под угрозой южных тропических лесах умеренного пояса Южной Америки. Каждая особь L. rosea производит меньше семян в результате снижения опыления и снижения способности пыльцевых зерен к прорастанию. В целом, это снижение репродуктивного успеха может, в свою очередь, привести размер популяции к демографическому вихрю, который может привести к исчезновению. Ни плотность нектара, ни плотность цветков не объясняют снижение уровня опыления L. rosea во фрагментах. Несмотря на более низкую влажность воздуха во фрагментах, одинаковое количество нектара с той же концентрацией сахара производят цветы, независимо от того, произрастают ли они в лесных остатках или сплошном лесу. Следовательно, цветы в равной степени привлекательны для опылителей, поскольку они предлагают одинаковую награду за услуги по опылению (Циммерман 19).88; Ратке 1992). С другой стороны, несмотря на тенденцию к снижению плотности цветения на фрагментах леса, даже на фрагменте с плотностью цветения, близкой к сплошному лесу, посещаемость была снижена, что позволяет предположить, что одна только плотность цветения не может объяснить такое сокращение. В сочетании со сниженной плотностью цветения обилие опылителей снижается на фрагментах леса. На самом деле, отношение цветы/опылители колеблется от 0,3 в сплошном лесу, предполагая, что каждый цветок может посещаться более чем одним опылителем, до 2,5 в менее посещаемых фрагментах, что означает, что на один доступный цветок приходится менее одного опылителя. . Опылители, кроме того, что обеспечивают меньше посещений, также несут пыльцу более низкого качества. Пыльца из фрагментов прорастает реже, чем пыльца из сплошных лесов. Следовательно, будет оплодотворено меньшее количество семязачатков, что объясняет меньшее количество семян.
Следовательно, цветы в равной степени привлекательны для опылителей, поскольку они предлагают одинаковую награду за услуги по опылению (Циммерман 19).88; Ратке 1992). С другой стороны, несмотря на тенденцию к снижению плотности цветения на фрагментах леса, даже на фрагменте с плотностью цветения, близкой к сплошному лесу, посещаемость была снижена, что позволяет предположить, что одна только плотность цветения не может объяснить такое сокращение. В сочетании со сниженной плотностью цветения обилие опылителей снижается на фрагментах леса. На самом деле, отношение цветы/опылители колеблется от 0,3 в сплошном лесу, предполагая, что каждый цветок может посещаться более чем одним опылителем, до 2,5 в менее посещаемых фрагментах, что означает, что на один доступный цветок приходится менее одного опылителя. . Опылители, кроме того, что обеспечивают меньше посещений, также несут пыльцу более низкого качества. Пыльца из фрагментов прорастает реже, чем пыльца из сплошных лесов. Следовательно, будет оплодотворено меньшее количество семязачатков, что объясняет меньшее количество семян. установить на лесных фрагментах. Lapageria rosea на фрагментах леса имеет более низкую гетерозиготность, чем в сплошном лесу (Henrıquez 2002), что может объяснить сниженную способность к прорастанию. Такая низкая гетерозиготность предполагает более высокий уровень инбридинга среди L. rosea из фрагментов леса. Поток пыльцы между несколькими особями, попавшими внутрь фрагмента, объясняет депрессию инбридинга (Prober and Brown 19).94), из-за более высоких показателей самообороны (Reusch, 2001) и более частых спариваний между близкими родственниками (Fenster, 1991). На самом деле поток пыльцы L. rosea ограничен радиусом 15 м от цветка-источника в цветочный район (Вальдивия, неопубликованные данные), что делает маловероятным поток пыльцы между сплошным лесом и фрагментами. Ни аборты, ни хищничество плодов не объясняют угнетенное завязывание семян в фрагментах леса. Сходные показатели абортов плодов указывают на то, что нет сильных различий в доступности ресурсов между фрагментарными и сплошными лесами (Stephenson 19).
установить на лесных фрагментах. Lapageria rosea на фрагментах леса имеет более низкую гетерозиготность, чем в сплошном лесу (Henrıquez 2002), что может объяснить сниженную способность к прорастанию. Такая низкая гетерозиготность предполагает более высокий уровень инбридинга среди L. rosea из фрагментов леса. Поток пыльцы между несколькими особями, попавшими внутрь фрагмента, объясняет депрессию инбридинга (Prober and Brown 19).94), из-за более высоких показателей самообороны (Reusch, 2001) и более частых спариваний между близкими родственниками (Fenster, 1991). На самом деле поток пыльцы L. rosea ограничен радиусом 15 м от цветка-источника в цветочный район (Вальдивия, неопубликованные данные), что делает маловероятным поток пыльцы между сплошным лесом и фрагментами. Ни аборты, ни хищничество плодов не объясняют угнетенное завязывание семян в фрагментах леса. Сходные показатели абортов плодов указывают на то, что нет сильных различий в доступности ресурсов между фрагментарными и сплошными лесами (Stephenson 19). 81), предполагая, что более низкий репродуктивный успех фрагментов является следствием взаимодействия между растением и опылителями. Наконец, учитывая, что S. sephaniodes и B. dahlbomii являются единственными колибри и шмелями, обитающими в тропических лесах умеренного пояса на юге Южной Америки, и они посещают большое количество растений в этой зоне, уменьшение их численности может привести к снижению опыления. обслуживает до 20% древесных видов тропических лесов умеренного пояса Южной Америки, например, Escallonia sp., Fuchsia magellania, Mitraria coccinea, Tristerix corymbosus, которые также произрастают в лесу Маулино (Aizen et al. 2002). Следовательно, большая часть растений, населяющих этот лес, может столкнуться со снижением репродуктивного успеха, как L. rosea.
Благодарности Мы благодарны CONAF и Forestal Millalemu за предоставление разрешений на работу в их штатах. Это исследование было поддержано FONDECYT 1981050 и 1010852 J.A. Симонетти. К.А. Анри Кес также благодарит докторскую стипендию и грант Te´rmino de Tesis от Conicyt, а также Departamento de Postgrado y Postı´ tulo Университета Чили за Beca Parcial de Tesis de Postgrado.
81), предполагая, что более низкий репродуктивный успех фрагментов является следствием взаимодействия между растением и опылителями. Наконец, учитывая, что S. sephaniodes и B. dahlbomii являются единственными колибри и шмелями, обитающими в тропических лесах умеренного пояса на юге Южной Америки, и они посещают большое количество растений в этой зоне, уменьшение их численности может привести к снижению опыления. обслуживает до 20% древесных видов тропических лесов умеренного пояса Южной Америки, например, Escallonia sp., Fuchsia magellania, Mitraria coccinea, Tristerix corymbosus, которые также произрастают в лесу Маулино (Aizen et al. 2002). Следовательно, большая часть растений, населяющих этот лес, может столкнуться со снижением репродуктивного успеха, как L. rosea.
Благодарности Мы благодарны CONAF и Forestal Millalemu за предоставление разрешений на работу в их штатах. Это исследование было поддержано FONDECYT 1981050 и 1010852 J.A. Симонетти. К.А. Анри Кес также благодарит докторскую стипендию и грант Te´rmino de Tesis от Conicyt, а также Departamento de Postgrado y Postı´ tulo Университета Чили за Beca Parcial de Tesis de Postgrado. Ссылки Айзен М.А. 1998. Фрагментация леса и воспроизводство растений: связь опыления. В: Брунс С., Мантелл С. и Трагард С. (ред.), Последние достижения в области биотехнологии для сохранения деревьев и управления ими. IFS, Стокгольм, стр. 22–37. Айзен М.А. и Фейнсингер П. 1994а. Фрагментация среды обитания, местные насекомые-опылители и дикие медоносные пчелы в аргентинском «Чако-Серрано». Экол. заявл. 4: 378–392. Айзен М.А. и Фейнсингер П. 1994b. Фрагментация среды обитания, опыление и воспроизводство растений в сухом лесу Чако, Аргентина. Экология 75: 330–351.
Айзен М.А., Васкес Д.П. и Смит-Рамирез С. 2002. История естественного и консервационного хозяйства мутуализма планта-животных дель боске темпладо де Судамерика австрал. Revista Chilena de Historia Natural 75: 79–97. Арместо Дж.Дж., Роззи Р. и Леон-Лобос П.М. 1996. Ecologı´ de los bosques chilenos: sı´ ntesis y proyecciones. В: Арместо Дж.Дж., Виллагран К. и Арройо М.К. (ред.), Ecologı´ de los Bosques Nativos Chilenos. Editorial Universitaria, Сантьяго, стр.
Ссылки Айзен М.А. 1998. Фрагментация леса и воспроизводство растений: связь опыления. В: Брунс С., Мантелл С. и Трагард С. (ред.), Последние достижения в области биотехнологии для сохранения деревьев и управления ими. IFS, Стокгольм, стр. 22–37. Айзен М.А. и Фейнсингер П. 1994а. Фрагментация среды обитания, местные насекомые-опылители и дикие медоносные пчелы в аргентинском «Чако-Серрано». Экол. заявл. 4: 378–392. Айзен М.А. и Фейнсингер П. 1994b. Фрагментация среды обитания, опыление и воспроизводство растений в сухом лесу Чако, Аргентина. Экология 75: 330–351.
Айзен М.А., Васкес Д.П. и Смит-Рамирез С. 2002. История естественного и консервационного хозяйства мутуализма планта-животных дель боске темпладо де Судамерика австрал. Revista Chilena de Historia Natural 75: 79–97. Арместо Дж.Дж., Роззи Р. и Леон-Лобос П.М. 1996. Ecologı´ de los bosques chilenos: sı´ ntesis y proyecciones. В: Арместо Дж.Дж., Виллагран К. и Арройо М.К. (ред.), Ecologı´ de los Bosques Nativos Chilenos. Editorial Universitaria, Сантьяго, стр. 405–421. Баррет С.Ч.Х. и Кон Дж. Р. 1991. Генетические и эволюционные последствия небольшого размера популяции растений: последствия для сохранения. В: Фальк Д.А. и Холсингер К.Е. (ред.), Генетика и сохранение редких растений. Оксфорд, издательство Оксфордского университета, стр. 3–30. Бустаманте Р.О. и Кастор С. 1998. Упадок находящейся под угрозой исчезновения экосистемы умеренного пояса: руиловый (Nothofagus alessandrii) лес в центральной части Чили. биодив. Консерв. 7: 1607–1626. Камарго J.L.C. и Капос В. 1995. Комплексные краевые эффекты на влажность почвы и микроклимат в лесах Центральной Амазонии. Дж. Троп. Экол. 11: 205–221. Эллстранд Н.К. и Элам Д.Р. 1993. Популяционная генетика небольшого размера популяции: значение для сохранения растений. Анну. Преподобный Экол. Сист. 24: 217–242. Фенстер С. Б. 1991. Поток генов у Chamaecrista fasciculata (Leguminosae) I. Распространение генов. Эволюция 45: 398–409. Газул Дж. и Маклиш М. 2001. Репродуктивная экология деревьев тропических лесов в вырубленных и фрагментированных местообитаниях в Таиланде и Коста-Рике.
405–421. Баррет С.Ч.Х. и Кон Дж. Р. 1991. Генетические и эволюционные последствия небольшого размера популяции растений: последствия для сохранения. В: Фальк Д.А. и Холсингер К.Е. (ред.), Генетика и сохранение редких растений. Оксфорд, издательство Оксфордского университета, стр. 3–30. Бустаманте Р.О. и Кастор С. 1998. Упадок находящейся под угрозой исчезновения экосистемы умеренного пояса: руиловый (Nothofagus alessandrii) лес в центральной части Чили. биодив. Консерв. 7: 1607–1626. Камарго J.L.C. и Капос В. 1995. Комплексные краевые эффекты на влажность почвы и микроклимат в лесах Центральной Амазонии. Дж. Троп. Экол. 11: 205–221. Эллстранд Н.К. и Элам Д.Р. 1993. Популяционная генетика небольшого размера популяции: значение для сохранения растений. Анну. Преподобный Экол. Сист. 24: 217–242. Фенстер С. Б. 1991. Поток генов у Chamaecrista fasciculata (Leguminosae) I. Распространение генов. Эволюция 45: 398–409. Газул Дж. и Маклиш М. 2001. Репродуктивная экология деревьев тропических лесов в вырубленных и фрагментированных местообитаниях в Таиланде и Коста-Рике.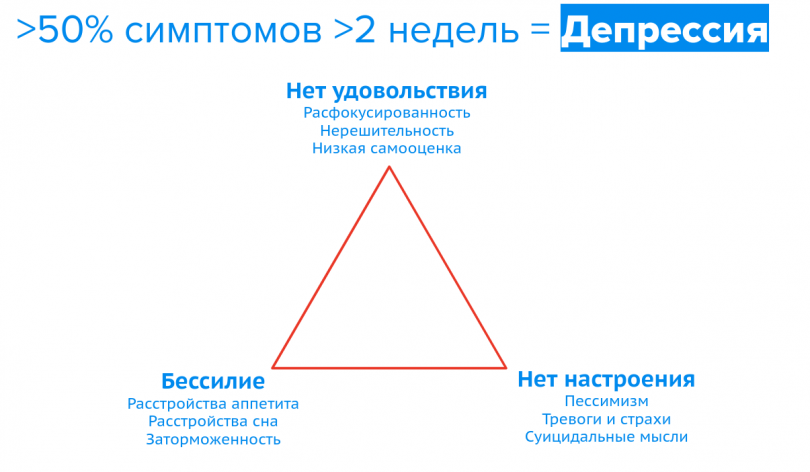 Завод Экол. 153: 335–345. Хаузер Т. П. и Зигизмунд Х. Р. 2000. Влияние инбридинга и аутбридинга на приспособленность пыльцы и выживаемость зигот у Silene nutans (Caryophyllaceae). Дж. Эвол. биол. 13: 446–454. Анри Кес C.A. 2002. El dilema de Lapageria rosea en bosques fragmentados: cantidad o calidad de la progenie?. Кандидат наук. Диссертация, Facultad de Ciencias. Университет Чили, Сантьяго, Чили. Human˜a A.M. и Риверос М. 1994. Биология размножения трепадоры Lapageria rosea R. et P. (Philesiaceae). Гаяна Ботаника 51: 49–55. Дженнерстен О. 1988. Опыление Dianthus deltoides (Caryophyllaceae): влияние фрагментации среды обитания на посещаемость и завязывание семян. Консерв. биол. 2: 359–366. Johannsson M.H., Gates M.J. и Stephenson A.G. 1998. Депрессия инбридинга влияет на производительность пыльцы Cucurbita texana. Дж. Эвол. биол. 11: 579–588. Капос В. 1989. Влияние изоляции на состояние воды в лесных массивах бразильской Амазонии. Дж. Троп. Экол. 5: 173–185. Кернс К.А. и Иноуэ Д.В. 1993.
Завод Экол. 153: 335–345. Хаузер Т. П. и Зигизмунд Х. Р. 2000. Влияние инбридинга и аутбридинга на приспособленность пыльцы и выживаемость зигот у Silene nutans (Caryophyllaceae). Дж. Эвол. биол. 13: 446–454. Анри Кес C.A. 2002. El dilema de Lapageria rosea en bosques fragmentados: cantidad o calidad de la progenie?. Кандидат наук. Диссертация, Facultad de Ciencias. Университет Чили, Сантьяго, Чили. Human˜a A.M. и Риверос М. 1994. Биология размножения трепадоры Lapageria rosea R. et P. (Philesiaceae). Гаяна Ботаника 51: 49–55. Дженнерстен О. 1988. Опыление Dianthus deltoides (Caryophyllaceae): влияние фрагментации среды обитания на посещаемость и завязывание семян. Консерв. биол. 2: 359–366. Johannsson M.H., Gates M.J. и Stephenson A.G. 1998. Депрессия инбридинга влияет на производительность пыльцы Cucurbita texana. Дж. Эвол. биол. 11: 579–588. Капос В. 1989. Влияние изоляции на состояние воды в лесных массивах бразильской Амазонии. Дж. Троп. Экол. 5: 173–185. Кернс К.А. и Иноуэ Д.В. 1993. Методы для биологов-опылителей. Университетское издательство Колорадо, Колорадо. Лопес-Каллеха М.В., Бозинович Ф. и дель Рио К. 1997. Влияние концентрации сахара на питание колибри и потребление энергии. Комп. Биохим. физ. А 118: 1291–1299. Мэтлак Г.Р. 1993. Изменчивость микросреды внутри и между лесными опушками на востоке США. биол. Консерв. 66: 185–194. Мурсия С. 1996. Фрагментация леса и опыление неотропических растений. В: Шелхас Дж. и Гринберг Р. (ред.), Лесные участки, тропические пейзажи. Island Press, Вашингтон, стр. 19.– 36. Pleasants J.M. 1983. Модели производства нектара Ipomopsis aggregata (Polemoniaceae). Являюсь. Дж. Бот. 70: 1468–1475. Пробер С.М. и Браун Х.Д. 1994. Сохранение травянистых белых самшитовых лесов: популяционная генетика и фрагментация Eucaliptus albens. Консерв. биол. 8: 1003–1013. Ramsey M. и Vaughton G. 1996. Депрессия инбридинга и наличие опылителей у частично самоопыляющегося многолетнего растения Blandfordia grandiflora (Liliaceae). Ойкос 76: 465–474.
Методы для биологов-опылителей. Университетское издательство Колорадо, Колорадо. Лопес-Каллеха М.В., Бозинович Ф. и дель Рио К. 1997. Влияние концентрации сахара на питание колибри и потребление энергии. Комп. Биохим. физ. А 118: 1291–1299. Мэтлак Г.Р. 1993. Изменчивость микросреды внутри и между лесными опушками на востоке США. биол. Консерв. 66: 185–194. Мурсия С. 1996. Фрагментация леса и опыление неотропических растений. В: Шелхас Дж. и Гринберг Р. (ред.), Лесные участки, тропические пейзажи. Island Press, Вашингтон, стр. 19.– 36. Pleasants J.M. 1983. Модели производства нектара Ipomopsis aggregata (Polemoniaceae). Являюсь. Дж. Бот. 70: 1468–1475. Пробер С.М. и Браун Х.Д. 1994. Сохранение травянистых белых самшитовых лесов: популяционная генетика и фрагментация Eucaliptus albens. Консерв. биол. 8: 1003–1013. Ramsey M. и Vaughton G. 1996. Депрессия инбридинга и наличие опылителей у частично самоопыляющегося многолетнего растения Blandfordia grandiflora (Liliaceae). Ойкос 76: 465–474. Rathcke BJ 1992. Распределение нектара, поведение опылителей и репродуктивный успех растений. В: Хантер М.Д., Огуши Т. и Прайс П.В. (ред.), Влияние распределения ресурсов на взаимодействие животных и растений. Academic Press, Нью-Йорк, стр. 113–138. Ройш Т.Б.Х. 2001. Последствия гейтоногамной саморегуляции приспособленности у клонального морского покрытосеменного растения (Zostera marina). Дж. Эвол. биол. 14: 129–138.
Сан-Мартин Дж. и Доносо С. 1996. Флористическая структура и влияние антропологии в эль-боске-маулино в Чили. В: Арместо Дж.Дж., Виллагран К. и Арройо М.К. (ред.), Ecologı´ de los Bosques Nativos de Chile. Editorial Universitaria, Сантьяго, стр. 153–168. Сих А. и Балтус М.С. 1987. Размер участка, поведение опылителей и ограничение количества опылителей у кошачьей мяты. Экология 68: 1679–1690. Смит-Рамирес С. 1993. Los picaflores y su recurso floral en el bosque templado de la Isla de Chiloe, Чили. Revista Chilena de Historia Natural 66: 65–73. Смит-Рамирез К. и Арместо Дж.
Rathcke BJ 1992. Распределение нектара, поведение опылителей и репродуктивный успех растений. В: Хантер М.Д., Огуши Т. и Прайс П.В. (ред.), Влияние распределения ресурсов на взаимодействие животных и растений. Academic Press, Нью-Йорк, стр. 113–138. Ройш Т.Б.Х. 2001. Последствия гейтоногамной саморегуляции приспособленности у клонального морского покрытосеменного растения (Zostera marina). Дж. Эвол. биол. 14: 129–138.
Сан-Мартин Дж. и Доносо С. 1996. Флористическая структура и влияние антропологии в эль-боске-маулино в Чили. В: Арместо Дж.Дж., Виллагран К. и Арройо М.К. (ред.), Ecologı´ de los Bosques Nativos de Chile. Editorial Universitaria, Сантьяго, стр. 153–168. Сих А. и Балтус М.С. 1987. Размер участка, поведение опылителей и ограничение количества опылителей у кошачьей мяты. Экология 68: 1679–1690. Смит-Рамирес С. 1993. Los picaflores y su recurso floral en el bosque templado de la Isla de Chiloe, Чили. Revista Chilena de Historia Natural 66: 65–73. Смит-Рамирез К. и Арместо Дж. Дж. 1998. Nectarivorı´ y polinizacion por aves en Embothrium coccineum (Proteaceae) en la Isla de Chiloe´, Чили. Revista Chilena de Historia Natural 71: 51–63. Soule´ ME 1986. Биология сохранения. Наука дефицита и разнообразия. Sinauer Associates, Suderland. Саутвик Э.Э., Лопер Г.М. и Садвик С.Е. 1981. Производство нектара, состав, энергия и привлекательность для опылителей весенних цветов западного Нью-Йорка. Являюсь. Дж. Бот. 68: 994–1002. Стефенсон А.Г. 1981. Аборт цветков и плодов: непосредственные причины и конечные функции. Анну. Преподобный Экол. Сист. 12: 253–279. Тамм С. и Гасс К.Л. 1986. Скорость потребления энергии и предпочтения концентрации нектара колибри. Экология 70: 20–23. Янг А., Бойл Т. и Браун Т. 1996. Популяционно-генетические последствия фрагментации среды обитания для растений. Тренд. Экол. Эвол. 11: 414–418. Циммерман М. 1988. Производство нектара, фенология цветения и стратегии опыления. В: Ловетт-Дауст Дж. и Ловетт-Дауст Л. (ред.), Репродуктивная экология растений, закономерности и стратегии.
Дж. 1998. Nectarivorı´ y polinizacion por aves en Embothrium coccineum (Proteaceae) en la Isla de Chiloe´, Чили. Revista Chilena de Historia Natural 71: 51–63. Soule´ ME 1986. Биология сохранения. Наука дефицита и разнообразия. Sinauer Associates, Suderland. Саутвик Э.Э., Лопер Г.М. и Садвик С.Е. 1981. Производство нектара, состав, энергия и привлекательность для опылителей весенних цветов западного Нью-Йорка. Являюсь. Дж. Бот. 68: 994–1002. Стефенсон А.Г. 1981. Аборт цветков и плодов: непосредственные причины и конечные функции. Анну. Преподобный Экол. Сист. 12: 253–279. Тамм С. и Гасс К.Л. 1986. Скорость потребления энергии и предпочтения концентрации нектара колибри. Экология 70: 20–23. Янг А., Бойл Т. и Браун Т. 1996. Популяционно-генетические последствия фрагментации среды обитания для растений. Тренд. Экол. Эвол. 11: 414–418. Циммерман М. 1988. Производство нектара, фенология цветения и стратегии опыления. В: Ловетт-Дауст Дж. и Ловетт-Дауст Л. (ред.), Репродуктивная экология растений, закономерности и стратегии.